Basic Embryology of Head and Neck
Embryological Development of Skeletal Structures of Head and Neck
Introduction
The structural development of the head and neck occurs between the third and eighth weeks of gestation. The 5 pairs of branchial arches, corresponding to the primitive vertebrae gill bars, that form on either side of the pharyngeal foregut on day 22 are the embryologic basis of all the differentiated structures of the head and neck. Each arch consists of 3 layers: an outer covering of ectoderm, an inner covering of endoderm, and a middle core of mesenchyme. These arches are separated further into external, ectoderm-lined pharyngeal clefts and internal, endoderm-lined pharyngeal pouches. This article summarizes the chronologic sequence of the embryologic development of the head and neck with respect to the branchial arches.
Week 1
After the sperm enters the oval cytoplasm in the uterine tube, the maternal and paternal chromosomes combine to form a zygote, completing fertilization. The ciliary action of the uterine tube transports the zygote to the uterus, during which it undergoes a series of mitotic divisions and transforms into the ball of small cells termed blastomeres. By the time they reach the uterus in approximately 3 days, the blastomeres become morula, a ball of 16 cells (after 4 mitotic divisions), and begin to form an internal cavity.1
Thus formed, the blastocyst consists of two layers of cells, an inner layer termed the embryoblast that becomes the embryo and an outer layer termed the trophoblasts that forms the embryonic part of the placenta. At approximately day 5, the blastocyst attaches to the endometrium in the posterior wall of the uterus, and at approximately day 6, the trophoblasts differentiate into syncytiotrophoblasts and cytotrophoblasts. The syncytiotrophoblasts then penetrate the endometrial epithelium and invade the underlying endometrial stroma; by the end of the first week, the superficial implantation of the blastocyst is complete.
Week 2
During the second week, a rapid proliferation and differentiation of the trophoblasts occurs. The decidual reaction of the endometrial tissues to accommodate the blastocyst implantation becomes accelerated during this week. At approximately day 9, the blastocyst is embedded completely beneath the endometrial epithelium, and spaces appear in the syncytiotrophoblasts that in 2 days form the lacunae network by fusion. The syncytiotrophoblast erosion into the endometrial blood vessels marks days 11 and 12.
Thus formed, uteroplacental circulation allows the maternal blood into the syncytiotrophoblast lacunae networks. Also in days 11 and 12, the primary yolk sac forms and part of the cytotrophoblast differentiates into extraembryonic mesoderm. The extraembryonic coelom derived from the extraembryonic mesodermal space eventually grows into the chorionic cavity.
By the end of week 2, the development of primary chorionic villi is observed, the primary yolk sac is replaced by the secondary yolk sac, and the amniotic cavity appears as a space between the cytotrophoblasts and the inner cell mass. The inner cell mass further differentiates into a bilaminar embryonic disc. The outer layer, the epiblast, is related to the amniotic cavity. The inner layer, the hypoblast, lies adjacent to the blastocyst cavity, and its localized thickening, the prochordal plate, becomes the future embryonic cranial region.
Week 3
The third week is marked by the formation of the trilaminar embryonic disc by the process of gastrulation; the inner layer is termed endoderm, the middle layer, mesoderm, and the outer layer, ectoderm. The embryo begins as a two-dimensional planar structure, and just prior to the third week, the stage is set for the development of the central nervous system in the area of ectoderm thickening known as the neural plate.
A midsagittal groove appears as a result of invagination of the ectoderm centrally and simultaneous elevation of ectodermal tissue alongside the groove to form the neural folds. These folds then fuse with each other in the mid line beginning at the junction of the future brain and spinal cord to form the neural tube. As the folds fuse, the neural tube separates from the overlying ectoderm from which it is derived.
A population of ectodermal cells adjacent to the neural fold and not included in the overlying surface (somatic) ectoderm gives rise to the formation of the neural crest. These neuroectodermal crest cells are believed to migrate widely throughout the developing embryo in a relatively cell-free enriched extracellular matrix and differentiate into a wide array of cell and tissue types, influenced by the local environment. Most connective and skeletal tissues of the cranium and face ultimately come from the derivatives of neural crest cells.
The first morphologic evidence of the optic primordia is seen as a thickened area with a shallow sulcus on the lateral forebrain of the neural tube in the region of the future diencephalon. During this week, the optic sulcus deepens and the surrounding walls form an invagination, which at first is in contact with the overlying ectoderm. Neural crest cells intervene between the optic vesicle and the overlying ectoderm and give rise to specialized neuroectodermal elements of the eye and adnexum.2
As the neural tube fuses cranially, closing off the anterior neuropore, a mesencephalic flexure occurs and the major divisions of the brain—prosencephalon (forebrain), mesencephalon (midbrain), and rhombencephalon (hindbrain)—can begin to be distinguished. By the end of the third week, further growth of the cephalic portion of the neural tube gives rise to the frontal nasal process and the branchial arches, the first stages of a recognizable face.3
Week 4
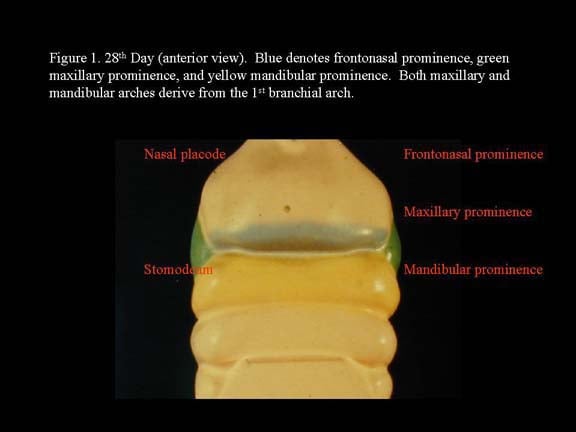
The recognizable face begins its development from 5 primordia that surround a central area of depression, the oral pit or the stomadeum. These primordia are the single cranially located frontal nasal process and two bilaterally located maxillary processes and mandibular processes. The maxillary and mandibular processes are derived from the first branchial arch. The first branchial arch, also termed the mandibular arch, forms the lateral wall and base of the stomadeum, the primitive mouth that appears as a slight depression of the ectoderm surface.4
The remaining branchial arches constitute the lateral and anterior walls of the primitive oropharynx, with the corresponding pharyngeal pouches between them. These processes or masses grow differentially, and by obliterating the ectodermal plates or grooves between them eventually give rise to the features of the face.4
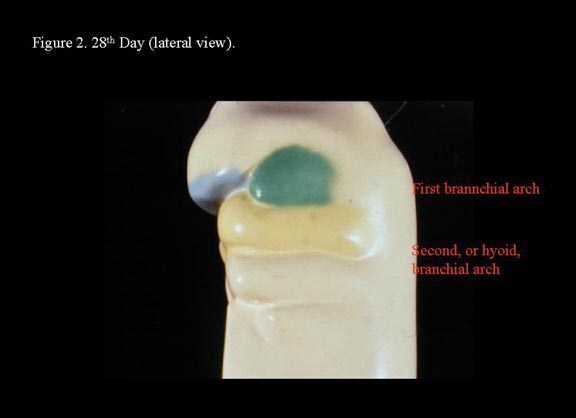
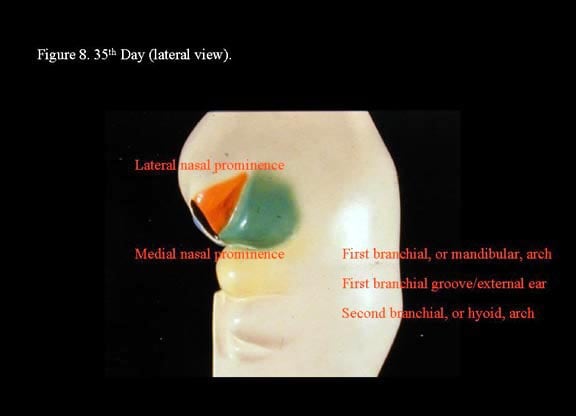
The mandibular processes are the first to merge with each other at the mid line and eventually give rise to the mandible and the lower part of the face and tongue. The dorsal end of the first arch cartilage, also termed Meckel cartilage, ossifies to form the malleus and incus of the middle ear. The dorsal end of the second arch cartilage, also termed Reichert cartilage, ossifies to form the stapes of the middle ear and the styloid process of the temporal bone. Small elevations at the dorsal ends of the first and second arches surrounding the first branchial groove fuse together to form the auricle of the external ear. The third branchial arch is not involved with the development of the ear.
The most common congenital anomaly of the branchial clefts is remnants of the second branchial cleft. The external component is found at the anterior junction of the middle and lower thirds of the sternocleidomastoid muscle and its track usually passes over the glossopharyngeal nerve and between the internal and external carotid arteries to the tonsillar fossa. The entire track and fossa must be excised. Cysts of the first branchial cleft are much less common but should be considered when excising a mass above the hyoid bone. Its course may involve the branches of the facial nerve. The third branchial cleft is rare but externally it may present at the same region as the second. It courses under the internal carotid artery.
During the fourth week, the primitive trisegmented brain further subdivides. The prosencephalon (forebrain) divides into the telencephalon (endbrain) with prominent lateral domes (cerebral hemispheres) and the diencephalon, which gives rise to the optic vesicles. The mesencephalon remains undivided and in the flexed cephalic flexure. The rhombencephalon divides into the mesencephalon (cerebellum, pons) and the myelencephalon (medulla). By the end of the fourth week, the fundamental organization of the future brain is clearly identifiable.4
The ectoderm overlying the optic vesicle of the developing eye thickens to form the lens placode. During this week, the optic vesicle deepens to form a goblet-shaped double-layered optic cup, as its stem narrows to form the optic stalk in continuity with the diencephalon. The frontal nasal process intervenes between the two lateral optic cups and at this stage the developing eyes are nearly 180° apart.
Head and neck embryology. 28th day (anterior view). Blue denotes frontonasal prominence, green maxillary prominence, and yellow mandibular prominence. Both maxillary and mandibular arches derive from the first branchial arch.
Head and neck embryology. 28th day (lateral view).
Week 5
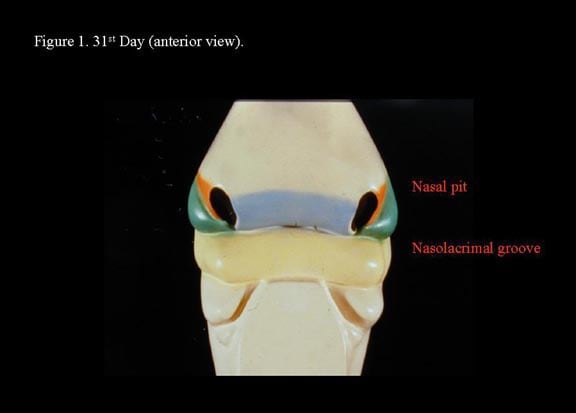
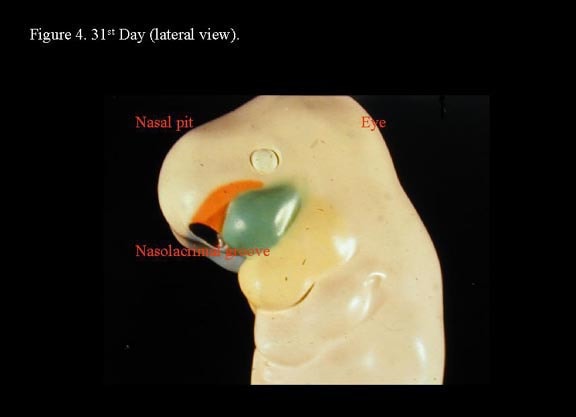
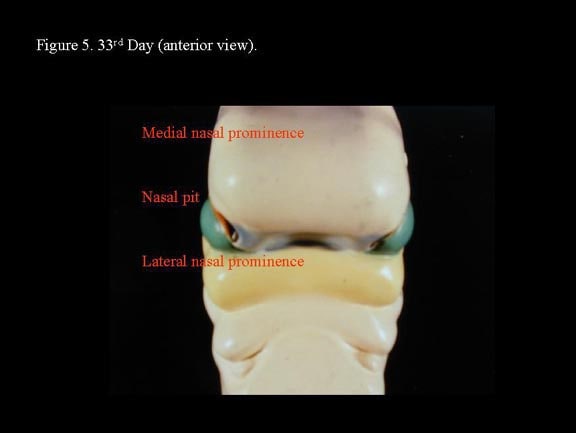
Toward the end of the fourth week, nasal placodes develop bilaterally at the inferolateral corners of the frontonasal process. As the region surrounding the nasal placodes develops into medial and lateral nasal processes, the deepening nasal placodes are transformed into nasal pits, in time forming the anterior nares. With continued ectodermal development, the nasal placodes become olfactory grooves that at this stage are in continuity with the developing oral cavity. The maxillary processes continue to enlarge, encroaching on the stomadeum, to form a primitive oral cavity.2
The first branchial arch gives rise to muscles of mastication (temporalis, masseter, lateral pterygoid, medial pterygoid). The mylohyoid, anterior belly of digastric, tensor tympani, and tensor veli palatini muscles also are derived from the first branchial arch. The second branchial arch develops into the muscles of facial expression.
The tongue is derived from the first through the fourth branchial arches. At the end of the fourth week, a median swelling, also termed median tongue bud, appears in the floor of the primitive pharynx just rostral to the foramen cecum. This is followed by a pair of lateral lingual swellings, also termed distal tongue buds, that arise from the proliferation of the ventromedial first branchial arches on each side of the median swelling. The distal tongue buds rapidly grow and fuse to form the anterior two thirds of the tongue, while the median tongue bud atrophies and forms no recognizable part of the adult tongue.
The posterior one third develops from the copula formed by the ventromedial part of the second branchial arches caudal to the foramen cecum and the hypopharyngeal eminence formed caudal to the copula from the ventromedial part of the third and fourth branchial arches. As the embryonic tongue further develops, the copula atrophies and disappears.5
The general mucosal sensory innervation of the anterior two thirds of the tongue, derived from the first arch, is from the lingual branch of the trigeminal nerve, the nerve of the first branchial arch. The chorda tympani branch of the facial nerve, the nerve of the second branchial arch, supplies the taste buds of the anterior two thirds of the tongue, except the vallate papillae.
The glossopharyngeal nerve, the nerve of the third branchial arch, supplies the taste buds in the vallate papillae and most of the posterior one third of the tongue mucosal surface. The superior laryngeal branch of the vagus nerve, the nerve of the fourth branchial arch, innervates a small patch of the tongue mucosal surface anterior to the epiglottis. All intrinsic and extrinsic tongue muscles, with the exception of the palatoglossus muscle, are supplied by the hypoglossal nerve. The palatoglossus muscle is supplied by the pharyngeal plexus fibers from the vagus nerve.
As the optic stalk narrows further to define the primitive optic nerve, retinal pigment appears in the external layer of the optic cup and the inner layer begins to differentiate into the neural elements of the retina. The primitive vitreous body develops in the intervening space as the lens placode invaginates toward the optic cup to form a lens vesicle, detaching itself from the overlying ectoderm.
As the features of the face form, the development of the embryonic skull begins. The mesenchyme surrounding the developing primitive brain condenses to form a mesenchymal capsule, the desmocranium. At approximately the fifth week, the base thickens, forming the cartilaginous cranial base through neuroectodermal condensation of the mesenchyme. In contrast, the calvaria develops by direct membranous ossification without a cartilaginous precursor later in the embryonic period.
Head and neck embryology. 31st day (anterior view).
Head and neck embryology. 31st day (lateral view).
Head and neck embryology. 33rd day (anterior view).
Head and neck embryology. 33rd day (lateral view).
Head and neck embryology. 35th day (anterior view).
Head and neck embryology. 35th day (lateral view).
Week 6
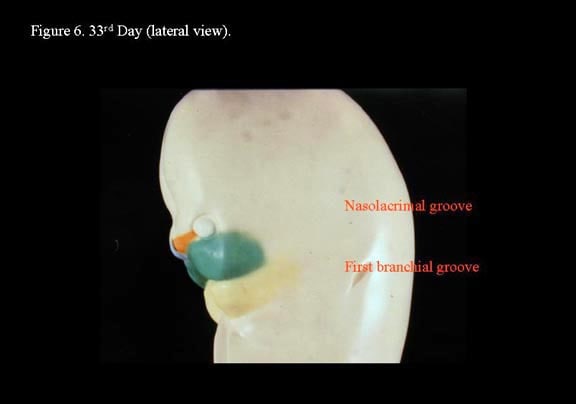
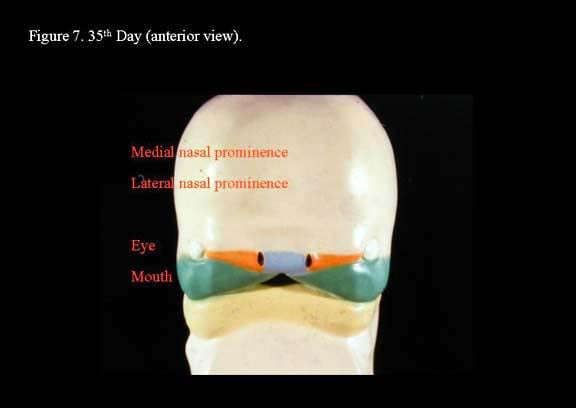
The medial nasal processes approach each other to form a single globular process that in time gives rise to the nasal tip, columella, prolabium, frenulum, and the primary palate. As this occurs, the frontonasal process collapses inward to form the nasal septum. Continued growth of the maxillary mass below the optic vesicles allows fusion with the lateral nasal process. As fusion occurs, a sinking troughlike epithelial groove demarcates the naso-optic furrow connecting the conjunctival lacrimal sac with the lateral nasal wall. The epithelial groove fuses as a solid epithelial cord and eventually canalizes later in fetal development to become the nasolacrimal duct.
Toward the end of the sixth week, the maxillary process fuses with the medial nasal fold of the globular process, forming a true nostril as it gives rise to the lateral lip element. However, posterior to this anterior fusion of the maxillary process to the nasal processes, the developing nasal floor is open to the oral cavity. Within the primitive stomadeum, lateral palatine processes develop from the medial edges of the maxillary process to give rise to the secondary palate. At this stage, the developing tongue nearly fills the oronasal cavity completely and reaches the nasal septum.4,2
Simultaneously, the eye continues its development as neural crest mesoectodermal cells at the lateral aspect of the lens differentiate into the early cornea and sclera. The cavity of the optic stalk is obliterated as nerve fibers from the retina grow back to reach the brain to form the optic nerve. The overlying surface ectoderm develops subtle horizontal folds, with mesenchymal core above and below the developing cornea representing the beginning of the upper and lower eyelids.
During this sixth week, the external ear develops from 6 mesenchymal swellings (hillocks) that surround the first branchial cleft. The first 3 hillocks arise from the mandibular arch (first branchial arch); the second 3, from the hyoid arch (second branchial arch). From ventral to dorsum, the hillocks of the first arch become the tragus, helix, and the cymba concha; the hillocks on the second arch become the antitragus, antihelix, and concha. The branchial cleft lengthens to form the primordium of the external auditory canal. As the face develops, the auricle gradually is repositioned to a more lateral cephalic position from the original location low on the side of the neck.
Week 8
By the eighth week, the face has acquired a more human appearance, with complete closure of the lower facial fissures or grooves. The primitive upper and lower jaws are formed with the complete fusion of the maxillary and mandibular processes. The upper lip and lower nasal regions are better defined. As it lengthens vertically, the frontonasal process continues to collapse horizontally, forming a transverse furrow at the nasal bridge. As the frontocranial region loses its overhang, the eyes begin to take a more medial position. The anterior rotation of the eyes continues throughout the remainder of the gestational period and well into childhood.6,2
The upper and lower lids continue to develop and assume a more almond shape as they begin to fuse with each other (separation of the lids not occurring until weeks 25-26). The lined ectodermal-derived epithelium between the fused eyelids and the cornea develops into the conjunctival sac, and from the superior-lateral angles, the lacrimal glands develop.
The development of the skull continues, with ossification centers arising within the mesenchyme (membranous ossification) adjacent to the developing brain to form the flat bones of the calvarial vault, the paired frontal and parietal bones. In contrast, the primordial elements of the chondrocranium begin to undergo endochondral ossification to form the bones of the basicranium (sphenoid, petrous part of temporal and occipital bones) and the nasal ethmoid significantly later in fetal life.
The sphenoethmoidal and sphenooccipital synchondroses and the septal cartilage persist to allow the forward growth of the cranial base and allow simultaneous inward rotation of the developing orbital complex. In the face, small nasal bones appear above the nasal capsule, the cartilaginous model. On the lateral surface of the nasal capsule, premaxillary and maxillary ossification centers appear.
Posterior-lateral to the developing maxillary bone, ossification centers arise within the zygomatic and squamoid portion of the temporal region. The mesenchyme encompassing Meckel cartilage undergoes membranous ossification to form the body of the mandible. In time, the Meckel cartilage disappears and persists as the incus and malleus of the middle ear. The onset of bone formation marks the end of the embryonic period and the developing fetus generally is much more resistant to teratogenic insults.7
Within this short period of 5 weeks, when the mother may be unaware of her own pregnancy, the face of her child has emerged. Any error that occurs in these tightly sequenced events of early facial embryogenesis can have significant consequences.
By whatever causative factor, failure of the lateral maxillary process to fuse with the central globular process is expected to result in a unilateral cleft of the upper lip. Similarly, the median cleft can be explained by incomplete merging of the median nasal processes, and an oblique facial cleft can be explained by persistence of the groove between the maxillary process and the lateral nasal process (medial canthal region to the ala of the nose).1
While the occurrence of some facial clefts7 can be understandable in terms of failure of fusion of the various primordial facial processes, others are not readily explainable in such terms. A cleft that begins lateral to the philtrum and extends to the mid portion of the lower eyelid is difficult to explain. Events prior to the normal process of fusion may lead to localized areas of tissue deficiency, possibly resulting in these atypical clefts.
Differences of opinion exist in the literature as to what fusion means in embryogenesis and how cell proliferation, cell differentiation, cell degeneration, and the extracellular matrix are involved in morphogenesis.
Neuromeric theory
Facial embryology was traditionally viewed as separate from the development of the brain. However, the intimate relation between facial and brain embryology has been made clearer through a better understanding of molecular genetics and induction. "The face predicts the brain," as written by DeMyer in 1964,3 has been replaced by the more contemporary "the brain predicts the face" of Carstens.2
The embryonic brain and spinal cord are divided into functional segments along the neuraxis that resemble a string of pearls. These units are termed neuromeres. The forebrain, or prosencephalon, contains 6 prosomeres. Prosomeres 1-3 and 4-6 constitute the primitive diencephalons and telencephalon, respectively. The midbrain, or mesencephalon, is derived from mesomeres 1 and 2. Rhombomeres 0-11 constitute the primitive hindbrain, or rhombencephalon. Each neuromere proceeding caudally from the hindbrain is flanked by a somitomere that subsequently differentiates into formed structures. Pertinent to facial anatomy, the first 7 somitomeres form the cranial base dorsally and the first 3 pharyngeal arches ventrally. The fourth through the sixth pharyngeal arches are formed by the ventral mesoderm of somitomeres 8-10.
The morphology of the face is produced by the assembly of embryonic fields according to genetic program. The German embryologist Wilhelm His originally described facial embryology as the fusion of structures that he termed processes. Fields represent true neuroangiosomes that move analogously to tectonic plates, while processes merely represent the topography of the embryonic face. Clefting is believed to result from the mechanical disruption of programmed field interactions. Better understanding of embryonic fields will hopefully lead to improved surgical strategies designed to address specific field pathology. The reader is referred to an excellent series of articles by Michael Carstens detailing the contemporary view of head and neck embryology relevant to the plastic surgeon involved in the treatment of congenital deformities.
Embryologic Development of DentitionThe primitive oral cavity, or stomodeum, is lined by stratified squamous epithelium called the oral ectoderm. Most of the connective tissue cells underlying the oral ectoderm are neural crest, ectomesenchyme, or neuroectoderm; these cells are thought to be responsible for the development of teeth. These cells develop from neural folds.8,9
At about 6 weeks (around 37 d) in utero, the ectoderm covering the oral cavity proliferates into a thick band of epithelium to form the horseshoe shape of the future alveolar process. This band is called the primary epithelium band, which quickly gives rise to subdivisions, the v estibular Lamina (lip furrow band) and dental lamina (maxillary and mandibular dental lamina).
The vestibular lamina proliferates into the ectomesenchyme, and its cells rapidly enlarge and degenerate to form a cleft that later becomes the vestibule between the cheek and alveolar process.
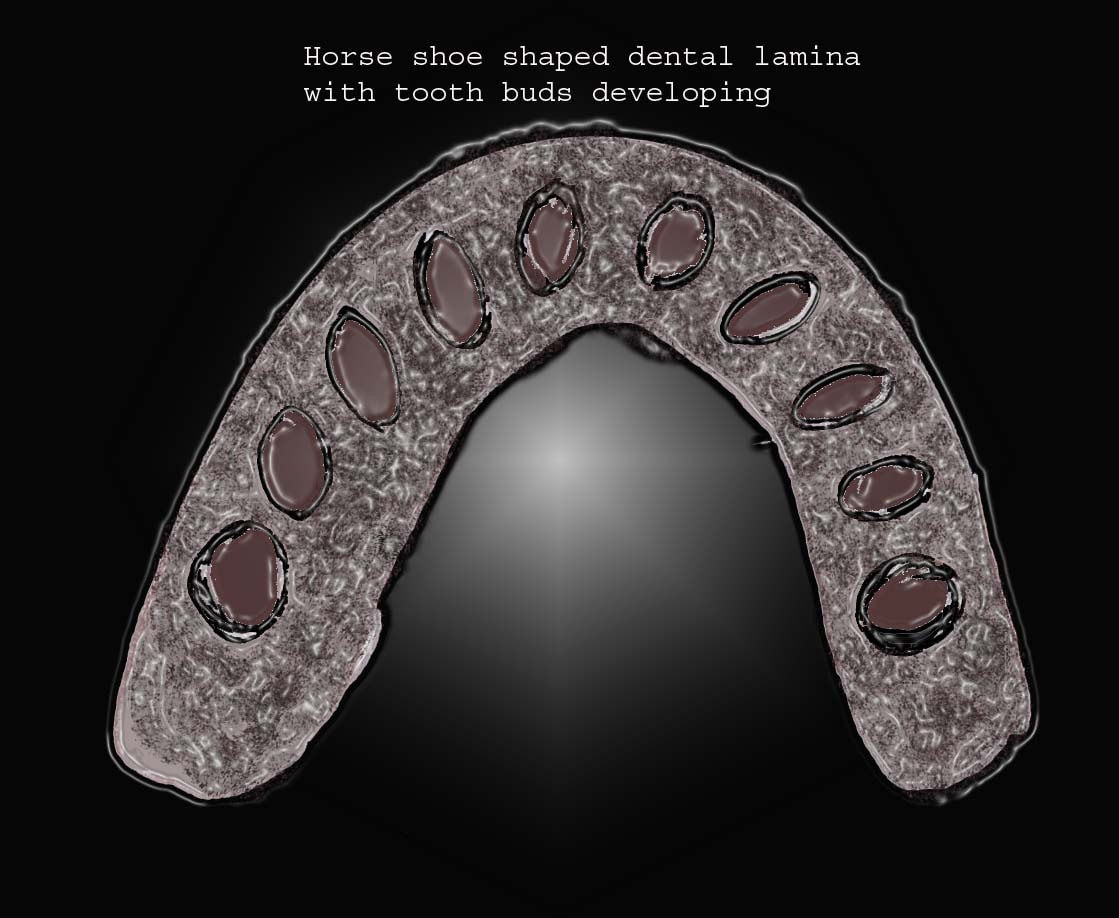
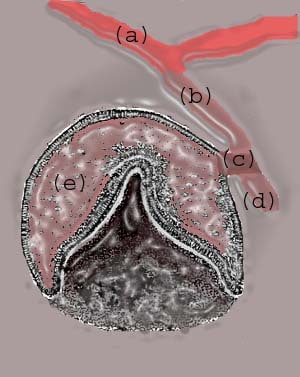
Localized proliferation occurs in the dental lamina, which leads to the formation of epithelial ingrowths into the ectomesenchyme, forming an ovoid structure called the placode at the future sites corresponding to 20 primary teeth. At this time, the mitotic activity is significantly lower than the corresponding ectomesenchyme, which suggests that part of the ingrowth is achieved by ectomesenchymal upward growth. The development of placode into tooth buds initiates from this stage. Dental lamina gives rise to 52 tooth buds (20 primary teeth by 6th and 8th prenatal weeks, 32 permanent teeth at later prenatal periods). The successional or permanent tooth buds develop lingually to the tooth buds of their deciduous predecessors; they develop from lingual extension of the dental lamina, which is called successional lamina.10
Placodes on the dental lamina for tooth development.
The activity of the dental lamina extends over a period of approximately 5 years and degenerates shortly after the initiation of tooth bud formation, but the lamina near the third molar continues to be active. Remnants of the dental lamina sometimes persist as epithelial pearls or islands within the jaw as well as in the gingiva.11
Variation in dental lamina can cause hyperdontia (more teeth), hypodontia (fewer teeth), or anodontia (complete or partial absence of more than 2 teeth).12
- Development of excess dental lamina mostly leads to increased tooth buds (hyperdontia). A single extra tooth commonly seen between the upper central incisors is called a mesiodens. Multiple supernumerary teeth are mostly associated with conditions such as cleft lip and palate, cleidocranial dysplasia, and Gardner syndrome. Supernumerary teeth may cause malocclusion and dentigerous cyst formation; they may also be impacted and cause resorption of root of the adjacent tooth.
- Deficient dental lamina leads to the absence of teeth. The most common tooth absent is the third molar, followed by the second premolar and the lateral incisors. Hypodontia is usually associated with microdontia (small teeth). Reduced alveolar development at the site is a common finding. Hypodontia is considered be genetically controlled, but environmental factors most likely influence the final result. The dental lamina is very sensitive to external stimuli that impair tooth bud development, such as trauma, infection, chemotherapeutic medications, endocrine disturbances, and severe intrauterine disturbances.
Some investigators believe that hypodontia is a normal variant, suggesting that humans are in an intermediate stage of dental evolution. The softer the diet, the lesser dentition needed for chewing; this may be why many people have missing third molars or impacted third molars. In patients with cleft involving the lip and palate, the dental lamina is fragmented and may result in hypodontia or, less frequently, supernumerary teeth in the cleft region. - Complete absence of teeth is known as anodontia. This is a rare condition usually seen in hereditary ectodermal dysplasia.
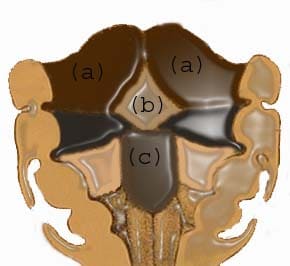
The tongue starts to develop at about 4 weeks. The tongue originates from the first, second, and third pharyngeal arches and forms the migration of muscles form the occipital myotomes.9 The anterior part arises from the first arch. A local proliferation of the mesenchyme gives rise to 3 swellings in the floor of the mouth; 2 lateral lingual swellings and a tuberculum impar arise from the mandibular process. The lateral lingual swellings enlarge rapidly and merge with each other; they overgrow the tuberculum impar to form the oral part of the tongue. Failure of the 2 lateral lingual swellings to merge causes cleft tongue or bifid tongue.13
A U-shaped sulcus develops in front of and on both sides of the oral part of the tongue. This allows the tongue to be free and highly mobile, except at the region of the lingual frenulum, where it remains attached. Disturbances during this stage cause tongue tie or ankyloglossia.14
The base of the tongue is formed mainly from the third pharyngeal arch. Initially, it is indicated by a midline elevation that appears behind the tuberculum impar, which is a large brachial eminence of the third and fourth arches. This eminence later becomes larger than the second arch, to become continuous with the body of the tongue. The site of union of body and base is delineated with a V-shaped sulcus called sulcus terminalis. Around the fifth and seventh weeks of gestation, the muscles forming the occipital myotomes migrate anteriorly. In the later stage of development, papillae differentiate on the dorsal mucosal covering the body of the tongue, whereas lymphatic tissue develops on the brachial part of the tongue.
Main masses that form the tongue: (a) lateral lingual swelling; (b) tuberculum impar; (c) hypobranchial eminence.
Formation and shaping of palatine shelves
At about 8 to 8.5 weeks of intrauterine life, the tongue helps shape the palatal bones. The lateral palatine shelves slide or roll over the body of the tongue. The process usually occurs from the combined action of the tongue movement and the action of the palatine shelves.8
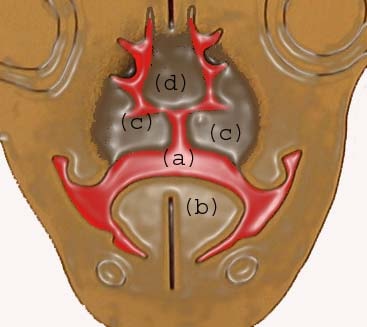
At 7 weeks: (a) communication between oral and nasal cavity; (b) tongue; (c) developing palatal shelf; (d) nasal septum.
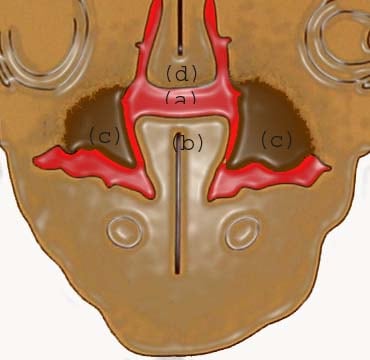
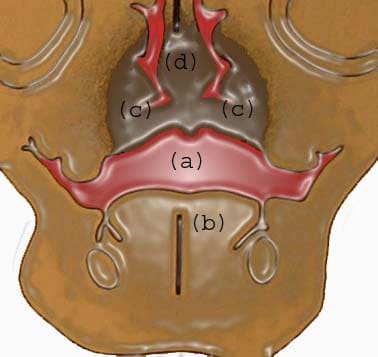
At 8 weeks: (a) communication between oral and nasal cavity; (b) tongue; (c) developing palatal shelf shaped by the musculature of the tongue; (d) nasal septum.
(a) Oral cavity; (b) tongue; (c) palatal shelves fused; (d) nasal septum.
Multimedia
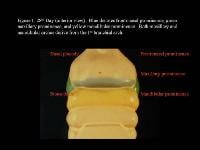 | Media file 1: Head and neck embryology. 28th day (anterior view). Blue denotes frontonasal prominence, green maxillary prominence, and yellow mandibular prominence. Both maxillary and mandibular arches derive from the first branchial arch. |
 | Media file 2: Head and neck embryology. 28th day (lateral view). |
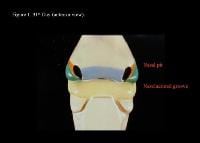 | Media file 3: Head and neck embryology. 31st day (anterior view). |
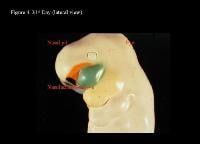 | Media file 4: Head and neck embryology. 31st day (lateral view). |
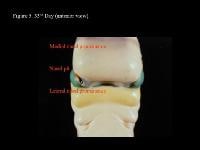 | Media file 5: Head and neck embryology. 33rd day (anterior view). |
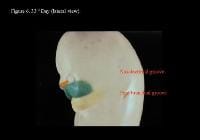 | Media file 6: Head and neck embryology. 33rd day (lateral view). |
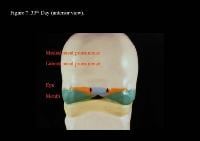 | Media file 7: Head and neck embryology. 35th day (anterior view). |
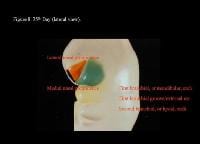 | Media file 8: Head and neck embryology. 35th day (lateral view). |
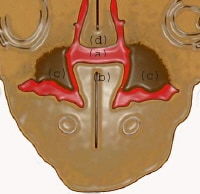 | Media file 9: At 7 weeks: (a) communication between oral and nasal cavity; (b) tongue; (c) developing palatal shelf; (d) nasal septum. |
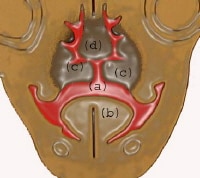 | Media file 10: At 8 weeks: (a) communication between oral and nasal cavity; (b) tongue; (c) developing palatal shelf shaped by the musculature of the tongue; (d) nasal septum. |
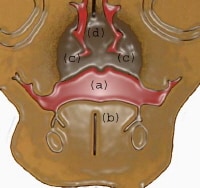 | Media file 11: (a) Oral cavity; (b) tongue; (c) palatal shelves fused; (d) nasal septum. |
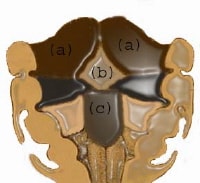 | Media file 12: Main masses that form the tongue: (a) lateral lingual swelling; (b) tuberculum impar; (c) hypobranchial eminence. |
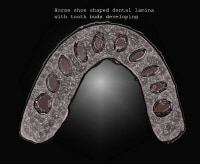 | Media file 13: Placodes on the dental lamina for tooth development. |
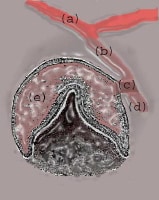 | Media file 14: (a) Oral epithelium; (b) dental lamina; (c) lateral lamina; (d) successional lamina; (e) tooth bud or enamel organ (future tooth). |
Keywords
head and neck embryology, head embryology, neck embryology, head development, neck development, fetal head, fetal neck, Development of teeth, development of dentition, tongue development